Scientists at Berkeley build a new Bay Bridge
Expanding the genome editing toolbox

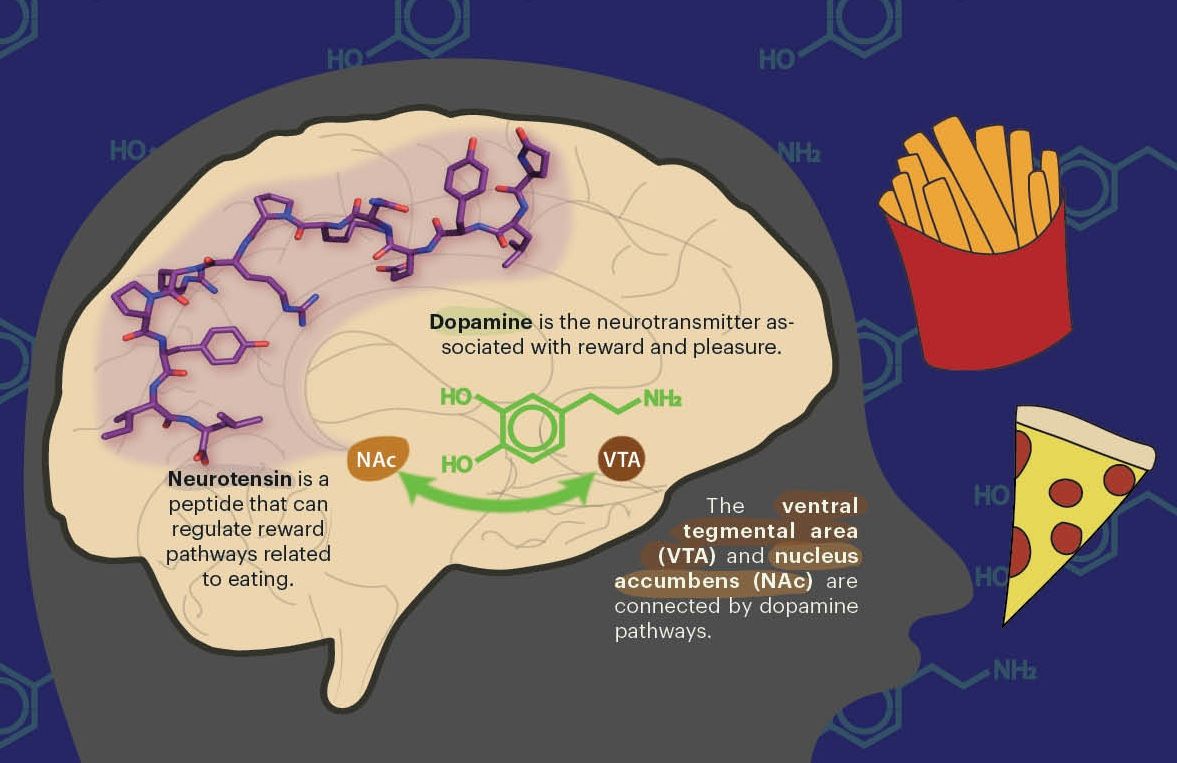
Early in 1998, biologists Andy Fire and Craig Mello published a paper with striking findings. By adding double-stranded RNA molecules into living cells, they were able to specifically and robustly inhibit genes of their choice. For much of the 20th century, RNA had been thought of as less important than DNA or proteins—the other molecules comprising the “central dogma” of molecular biology. Fire and Mello’s technology, now known as RNA interference (RNAi), marked a crucial moment in RNA’s journey to greater recognition by showcasing how it could be used programmably.
Just over a decade later, researchers similarly reprogrammed the bacterial immune system CRISPR to cut DNA in a sequence-specific manner. CRISPR consists of an RNA molecule called a guide that directs it to the correct sequence and a nuclease protein that then cleaves the DNA. By changing the sequence of the guide RNA, the complex could be directed to cut at different DNA bases. Thus, by exploiting cells’ natural DNA repair mechanisms, CRISPR enabled specific edits to small stretches of DNA. As powerful as the technology has become in the intervening years, it still lacks the ability to insert large DNA sequences—a tantalizing problem that has motivated researchers for decades. Fortunately, Patrick Hsu’s lab at UC Berkeley and the Arc Institute recently discovered a system capable of doing just that.
In an effort co-led by senior scientist Matt Durrant and PhD student Nick Perry, Hsu’s team characterized a system capable of producing large-scale DNA rearrangements with programmability similar to RNAi or CRISPR. The system consists of a recombinase enzyme that carries out the changes and a “bridge RNA” that can be altered to allow the insertion of custom sequences in a site-specific manner. While the bridge RNA system is brand new as a bioengineering tool, it has the potential to impact biological research and therapeutic development tremendously, pushing the genome engineering field to new heights.
Genetic detective work
Recombinases are proteins that can swap out two different DNA sequences, though typically not in a programmable manner. While biologists have long utilized recombinases to perform insertions, the enzymes have traditionally been limited to very specific insertion sites, rendering most spots in any given genome inaccessible. Hoping to find novel genome editing tools in nature, Hsu’s team began searching for enzymes with similar properties that might also be RNA-guided. They began by looking at the DNA sequences of transposons—small “jumping genes” that encode the components necessary to “cut and paste” themselves throughout their hosts’ genomes. Before starting starting this search, Durrant and other lab members had been studying TnpBs—small RNA-guided nucleases similar to Cas9 but encoded by transposons.
Given that transposons typically encode recombinases or transposases capable of large DNA insertions, the mobile elements offered a promising starting place. In the course of this detective work, Durrant found a few clues that led him to a special class of transposons known as IS110s. Earlier research had demonstrated that these transposons were active in their host organisms but had notably failed to replicate their activity in test tubes. Something was missing. As Durrant puts it, “sometimes just the fact that something’s not working—the absence of evidence—is evidence.” Moreover, Perry notes that IS110s’ site-specific recombination mechanism was “fairly unusual for mobile elements” and might also be explained by the system’s missing component.
When Durrant looked closer at IS110 genomic loci, he noticed that the ends of the transposons—which don’t code for the recombinase—were abnormally long. Putting a lot of stock in these elements still seemed a bit risky, but if the noncoding ends produced a guide RNA, the potential payoff was huge. Durrant and Perry began working together in earnest, with Durrant providing computational analyses and Perry testing the system in the lab. Both researchers emphasized the importance of collaboration to the project’s success, with Durrant stating that “a big feature of all of this was that we worked very well together, and that was a big reason why we were able to move so quickly.”
Seeking a representative sequence for their biochemical analyses, Perry and Durrant zeroed in on the IS621 transposon, an IS110 family member found in the ubiquitous bacterium E. coli. Their first challenge was to confirm that there really was an RNA molecule encoded by the element. To do this, they used a common technique called RNA sequencing, which allowed them to identify any parts of the element that produced a noncoding RNA. After numerous unsuccessful attempts, Perry finally created a biological construct that, upon sequencing, showed an RNA at the right end of the transposon sequence. “The signal was a straight-up peak,” he remembers. “It was shocking. All of this happened in the span of two to three weeks, where we saw this signal computationally and verified it in the lab.” He and Durrant quickly confirmed that the IS621 recombinase did in fact bind this RNA, indicating that it likely was the missing component of the system. Now, they just had to determine exactly how it worked. Given the site-specificity of IS110 elements, Durrant reasoned that that if the RNA served as a guide, it would have a region matching the DNA sequence being targeted for the recombination event. To assess this possibility, he turned to a technique called covariation analysis, which allowed him search thousands of IS110 sequences for pairs of nucleotides that frequently matched each other. He found not one but two covariation hotspots in the RNA sequence, ultimately deducing that one indicated the region that bound the insertion target site, while the other bound the “donor” DNA to be inserted: the transposon itself. Durrant quickly realized that the system was “almost certainly RNA-guided and programmable,” though in a somewhat unexpected place.
“I was in the middle of a move, and I was working in my parents’ pantry,” he says, pulling up a picture of his desk and computer amid shelves of soup cans and paper towels. “I was so happy I couldn’t sleep for a week, but it was awesome."
Using other computational tools, he predicted the average structure of more than 100 1S110 RNAs, revealing that the donor-binding and target-binding sequences each occupied a characteristic loop in the molecule. Realizing that the guide RNA directly connected the two DNA sequences to be exchanged, Hsu’s team decided to call it a “bridge RNA.” As Perry began his next set of experiments with the system, he began sending samples to structural biologist Hiroshi Nishimasu’s group at the University of Tokyo. Nishimasu’s team collected high-resolution structures of the bridge RNA in action, showcasing precisely how the bridge RNA system achieved targeted recombination. Perry reiterates the importance of collaboration to the project, stating that “these proteins are not easy to work with, and their experience was very valuable.”
Engineering a programmable system
Having uncovered the missing component in IS110 recombination, Hsu’s team now had to demonstrate that the bridge RNA could be reprogrammed like RNAi and CRISPR. They started with a proof-of-principle experiment in a test tube, creating a series of bridge RNAs matching seven different spots on a circular target DNA molecule that they placed inside the workhorse research bacterium E. coli. Remarkably, they were able to successfully insert the IS621 donor sequence into all of them. With these data in hand, they turned to the more difficult problem of demonstrating programmable recombination in a biologically relevant context. To do this, they set up a similar experiment, creating bridge RNAs designed to insert at specific points in the E. coli genome.
To demonstrate that the donor DNA could also be programmably altered, they added a nearly 5000-nucleotide molecule to their E. coli set up and altered the other bridge RNA loop to target it. They observed recombination of the foreign DNA into the E. coli genome. While they observed a small number of “off-target” integrations at sites not directly targeted by the bridge RNA, the intended site was always the most frequently targeted. The system wasn’t perfect, but it was fully programmable and worked in living cells with barely any engineering from Hsu’s team. In a final set of experiments, the team showed that the bridge RNA system could also be programmed to execute other large-scale DNA arrangements. With this demonstration that the bridge RNA system could produce sequence-specific excisions and inversions, they had recapitulated the most sought-after features of non-programmable recombinases.
A new era of genome editing
The whole bridge RNA apparatus is quite compact, a feature that Durrant says may be due to the evolutionary pressure acting on transposons themselves. “Maybe these are smaller in part because mobile elements survive better when they’re smaller,” he speculates. This limited size makes the tool even more tantalizing for therapeutic applications, which require editing tools to be packaged for delivery into cells. It remains to be seen, however, whether bridge RNAs’ efficiency in bacteria will translate to mammalian cells. Fortunately, its mechanistic novelty makes the bridge RNA an intriguing basis for therapeutics in cell types where it exhibits even modest activity. Durrant predicts that in its fully optimized form, the bridge RNA has the potential to be much more impactful than other fully optimized genome editing tools. “I think basic Cas9 is basically at the limit, so now people are adding new domains to create new tools and then optimizing those,” he says. “The bridge RNA has a new chemistry altogether and as we engineer more specificity and make it efficient in human cells, it will open up a whole new landscape of edits.”
The possibilities are endless, and Perry believes that the technology will ultimately incite a paradigm shift from genome editing to genome design. “What we’re trying to do here is not edit the words in a paragraph or change the paragraphs on a page but rewrite entire pages of chapters of a book, with the book being the genome.” Of course, Durrant and Perry also believe that the bridge RNA system will make a splash outside of human therapeutics.
Perry has “a whole page worth of lists of ideas of applications,” while Durrant sees the bridge RNA as a useful tool for insect and plant genome editing, diagnostic tools, and even common biology lab workflows such as molecular cloning. For now, their goal is to make the bridge RNA as accessible as possible. Perry believes that system will be useful to researchers across molecular biology if it’s readily available and easy to use. “Problems can be solved by manipulating the genome or modifying your protein of interest or modifying your cell,” he says. “You can programmably remove, rearrange, or insert whatever you need to understand how the genome works or how your particular disease works.” Indeed, the Arc Institute has already created an online tool that researchers can use to generate custom bridge RNAs for their own research needs. If more biologists take up the call and study the bridge RNA system, its development will undoubtedly be accelerated. “I just want to see what people are going to do with it,” Durrant says.